A rekombináns fehérjetechnológia a modern biotechnológia sarokkövévé vált, lehetővé téve a fehérjék nagymértékű előállítását kutatási, ipari és terápiás alkalmazásokhoz. Az optimális fehérje expressziós rendszer kiválasztása kulcsfontosságú a hatékony rekombináns fehérjetermelés eléréséhez, a helyes feltekeredéssel, poszttranszlációs módosításokkal és biológiai aktivitással. A főbb rekombináns fehérje expressziós rendszerek osztályozásának és jellemzőinek megértése értékes útmutatást nyújt a kísérleti tervezéshez és a fehérjetisztításhoz.
A Rekombináns Fehérjetermelés Alapjai és Lépései
A rekombináns fehérjetermelés egy olyan eljárás, amelynek során idegen géneket expresszálnak és célfehérjéket állítanak elő gazdasejtekben géntechnológiával. Ezt a technológiát általában olyan fehérjék előállítására használják, amelyeket nehéz beszerezni, vagy amelyek természetes állapotban nem állíthatók elő hatékonyan, például különféle fehérjék orvosi, tudományos, ipari és biotechnológiai alkalmazásokhoz. A géntechnológia alapján exogén géneket expresszálnak és célfehérjéket állítanak elő a gazdasejtek transzkripciós és transzlációs mechanizmusainak felhasználásával. A lényege, hogy egy specifikus fehérjét kódoló gént (általában egy idegen gént) bejuttatnak a gazdasejtbe, és a célfehérjét a sejt természetes bioszintézisének folyamatán keresztül állítják elő.
A rekombináns fehérje előállításának kísérleti lépései általában a következő főbb lépéseket foglalják magukban. Az egyes lépések sikeres befejezése kulcsfontosságú a végső, kiváló minőségű rekombináns fehérje előállításához:
Génklónozás és Vektorépítés
Ez a fázis a célgén azonosításával és előkészítésével kezdődik. A célfehérjék kódoló génjeit forrásorganizmusokból (például emberekből, baktériumokból, növényekből stb.) vonták ki. A célgént gyakran PCR-rel (polimeráz láncreakcióval) amplifikálják. Ezután a célgént egy expressziós vektorba (például plazmidba) klónozzák. Az expressziós vektorok általában promótereket, tag szekvenciákat, szelekciós markereket stb. tartalmaznak, amelyek képesek idegen gének expresszióját irányítani a gazdasejtekben. A létrehozott vektort emésztik és szekvenálják, hogy biztosítsák a célgén helyes beépülését a vektorba.
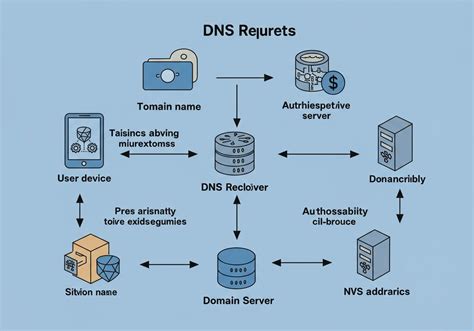
Gazdasejtek Kiválasztása és Transzformációja/Transzfekciója
A célfehérje jellegétől és a termelési igényektől függően válassza ki a megfelelő gazdasejteket. Gyakran használt gazdasejtek közé tartoznak az Escherichia coli (E. coli), az élesztő, a rovarsejtek, az emlőssejtek stb. Prokarióta transzformáció (prokarióta gazdasejtek, például E. coli esetén) során az expressziós vektorokat hősokkkal, kémiai transzformációval vagy elektroporációval juttatják be a gazdasejtekbe. Eukarióta gazdaszervezetek, például emlőssejtek esetében, transzfekcióval juttatják be a vektort kémiai reagensekkel (például liposzómákkal) vagy elektroporációs technikákkal. A sikeresen transzformált vagy transzfektált sejteket antibiotikum-szűréssel szelektálják (például a vektorban található antibiotikum-rezisztencia gének felhasználásával).
Fehérje Expresszió
Ha a felhasznált gazdasejt szabályozható expressziós rendszerrel rendelkezik (például az E. coli T7 rendszerével), a célfehérje expresszióját induktorok (például IPTG) hozzáadásával aktiválják. Az expressziós feltételek optimalizálása során a célfehérje expressziós szintjét és feltekeredési állapotát a tenyésztési hőmérséklet, a táptalaj összetétele, az indukciós idő és egyéb feltételek beállításával optimalizálják. A kifejezés ellenőrzésére SDS-PAGE, Western blot és más módszereket alkalmaznak a célfehérje expressziójának előzetes ellenőrzésére, hogy biztosítsák annak expressziójának mennyiségét és minőségét.
Sejtgyűjtés és Lízis
A gazdasejteket centrifugálással választják el a táptalajtól, és általában lefagyasztják, vagy közvetlenül lízishez használják. A sejteket fizikai módszerekkel (például ultrahangos roncsolás, nagynyomású homogenizálás) vagy kémiai módszerekkel (például lízispuffer, liáz stb.) roncsolják a bennük lévő fehérjék felszabadítása érdekében. A lízisoldatot SDS-PAGE-el elemzik a fehérje felszabadulásának megerősítésére.
Fehérjetisztítás
A célfehérje jellemzői alapján választják ki a megfelelő tisztítási módszert. Az affinitáskromatográfia a célfehérje szelektív tisztítását teszi lehetővé az affinitási jelölés (például His-jelölés) affinitásoszlophoz való kötésével. Az ioncserélő kromatográfia a különböző fehérjéket töltési jellemzőik alapján választja szét. A gélszűréses kromatográfia a fehérje molekulatömegén alapuló elválasztást biztosítja. Tisztítás utáni ellenőrzés során SDS-PAGE, Western blot és más technikákat alkalmaznak a célfehérje tisztaságának és molekulatömegének megerősítésére.
Fehérjeaktivitás-ellenőrzés és Jellemzés
Biológiai módszerekkel ellenőrzik, hogy a fehérje rendelkezik-e a várt biológiai aktivitással, például enzimaktivitási vizsgálattal, antitestkötési vizsgálattal stb. A fehérje szerkezetét tovább elemzik tömegspektrometriával, mágneses magrezonanciával (NMR) vagy röntgenkrisztallográfiával. A fehérje stabilitását termikus stabilitással, pH-stabilitási vizsgálattal és egyéb kísérletekkel értékelik.
SEJTBIOLÓGIA - SEJTEK SZERKEZETE
Fehérjetárolás és -felhasználás
A fehérje jellemzőitől függően választják ki a megfelelő tárolási körülményeket (például krioprezerváció, fagyasztva szárítás stb.) a fehérje stabilitásának biztosítása érdekében. A tisztított rekombináns fehérje különféle alkalmazásokban használható, például tudományos kutatásban, vakcinafejlesztésben, enzimatikus alkalmazásokban, gyógyszergyártásban stb.
Rekombináns Fehérje Expressziós Rendszerek
Egy rekombináns fehérjét úgy állítanak elő, hogy a kívánt fehérjét kódoló gént bejuttatnak egy gazdaszervezetbe, amely képes azt átírni és transzlálni. A gazdaszervezet sejtes gépezete ezután szintetizálja a célfehérjét, amely lehet intracelluláris, szekretált vagy membránhoz kötött. A modern fehérjeexpressziós rendszereket nagyjából négy kategóriába sorolják: prokarióta rendszerek, élesztőrendszerek, rovarsejt-rendszerek és emlőssejt-rendszerek. Mindegyiknek egyedi előnyei, korlátai és megfelelő alkalmazásai vannak a fehérje komplexitásától, a hozamkövetelményektől és a poszttranszlációs igényektől függően.
Prokarióta Expressziós Rendszerek: E. coli
Az Escherichia coli továbbra is a legszélesebb körben használt gazdaszervezet rekombináns fehérjék előállításához egyszerűsége, alacsony költsége és gyors növekedése miatt. Lehetővé teszi a magas szintű fehérjeexpressziót erős promóterek és hatékony vektorrendszerek használatával. Ezenkívül a genetikai manipuláció és a skálázhatóság egyszerűsége ideálissá teszi enzimek, antitestfragmensek és ipari fehérjék előállítására.
A fehérjék biotechnológiai előállítása előnyösen olyan mikroorganizmusok segítségével történik, amelyek könnyen tenyészthetők, és a termelt fehérjék egyszerű módon kinyerhetők. Erre a célra alkalmas mikroorganizmus például a baktériumok közül a gramnegatív Escherichia coli. Az E. coliban a természetes fehérjék transzlációfüggő N-terminális metionincsoportja legtöbbször proteázok révén nagyrészt lehasad. Idegen fehérjék esetén azonban az első metionincsoportnak ez a lehasadása többnyire csak részleges. A rekombináns fehérjék prokarióta szervezetekben történő előállításánál ugyanis szükséges egy olyan DNS-szekvencia klónozása, amely a tulajdonképpeni DNS-szekvencia előtt kezdő kodonként egy AUG kodont tartalmaz. Ennek következtében prokariótákban, mint például E. coliban egy olyan rekombináns fehérje fejeződik ki, amely az első aminosav helyén (1-es pozícióban) metioninrészt tartalmaz. Sok esetben azonban az szükséges, hogy az 1-es pozícióban metioninmentes rekombináns fehérjéket állítsanak elő.
Az E. coli alkalmazása széleskörű, beleértve az orvosi vegytani kutatásokat is. Az Orvosi Vegytani Intézetben rekombináns DNS sokszorosítása E. coli baktériumokba vagy emlős sejtekbe való bejuttatás céljából történik, valamint rekombináns fehérjék, illetve ezek mutánsainak kifejezése E. coli alapú expressziós rendszerben a fehérje biológiai funkciójának in vitro vizsgálatára. Az Élettani Intézetben rekombináns fehérjék, illetve azok pontmutánsainak előállítása történik E. coli alapú expressziós rendszerben a fehérje biológiai funkciójának vizsgálatára. A Laboratóriumi Medicina Intézetben plazmidok előállítása E. coli tenyészetek és szintetikus DNS felhasználásával történik, mutagenizálás, heterológ expresszió, rekombináns fehérjék előállítása, funkcionális vizsgálatok mellett. A Humángenetikai Tanszéken (új nevén Orvosi Genetikai Tanszék) is E. coli tenyészeteket használnak plazmidok előállítására rekombináns DNS technológia segítségével. A Biofizikai és Sejtbiológiai Intézetben kompetens baktérium törzsek előállítása, baktériumok transzformációja genetikailag módosított örökítőanyaggal történik.
Francia tudósok azt vizsgálták, milyen módon befolyásolhatja az E. coli a gazdaszervezet étkezési szokásait. Állatkísérletek során azt figyelték meg, hogy az E. coli baktérium a tápanyagellátás által kiváltott szaporodási fázis után fehérjéket választ ki, melyek csökkentik az egerek és patkányok étvágyát. Sergueï Fetissov (University of Rouen, Franciaország) és kollégái azt feltételezik, hogy a baktériumok részt vesznek az étvágy szabályozásában, mégpedig úgy, hogy a táplálékfelvétel után nem sokkal egy fehérje termelődésén keresztül a gazdaszervezet bélfal sejtjeit jóllakottságot jelző hormon termelésére késztetik, amelyek a véráram útján az agyba kerülve speciális agysejteket aktiválnak. Táplálékhiány esetén e jelek kimaradása hozzájárul az éhségérzet keletkezéséhez. A kutatók vizsgálatai az E. coli baktériumra fókuszáltak. Fetissov és munkatársai a bélbaktériumok növekedési feltételeit szimulálták, a tenyészeteket szakaszosan látták el tápanyagokkal. Minden „etetést” egy 20 perces gyors sejtszaporodási fázis követett, amely után a baktériumok a fent említett, bélfalsejt stimuláló fehérjéket állították elő. Amennyiben ezeket a fehérjéket egerekbe vagy patkányokba injektálták, a kísérleti állatok bélsejtjei fokozottan termelték a jóllakottság jelző hormonokat, ezáltal pedig csökkent a táplálékfelvételük. A tápoldat hozzáadása előtt meglévő bakteriális fehérjék nem váltottak ki ilyen hatást. Egy, az etetés után termelt E. coli fehérje erős kémiai hasonlóságot mutat az α-MSH jóllakottság jelző hormonnal. A baktérium által termelt fehérje az állatok vérében is kimutatható, és ez aktiválja az étvágycsökkentő neuronokat a hipotalamuszban. A kutatók feltételezése szerint az E. coli baktériumon kívül valószínű, hogy más bélbaktériumok is részt vesznek az étvágy szabályozásában.
SEJTBIOLÓGIA - SEJTEK SZERKEZETE
Élesztő Expressziós Rendszerek
Az élesztő, különösen a Saccharomyces cerevisiae (sütőélesztő), gyakran használt eukarióta gazdaszervezet komplex rekombináns fehérjék előállítására, amelyek glikozilációt, hajtogatást és komplex módosítást igényelnek. Az élesztőrendszerek ötvözik az E. coli könnyű kezelhetőségét az eukarióta poszttranszlációs módosítások képességével. A Pichia pastoris egy másik népszerű élesztőfaj, amelyet magas hozamú fehérjetermelésre használnak, különösen szekretált fehérjék esetén.
Az élesztők közül a Saccharomyces cerevisiae alkalmas mikroorganizmus a fehérjék biotechnológiai előállítására. Azonban autentikus idegen gének kifejezése az ilyen mikroorganizmusokban mégis gyakran hátrányokkal jár. A Pichia pastoris nem patogén gomba, nem ismert patogenitása állatra, növényre vagy egyéb élőlényre. Rekombináns skorpiótoxintermelés kizárólag zárt rendszerben történik. A termelt toxinok 30-40 aminosavból álló peptidek. A peptidek szobahőmérsékleten viszonylag gyorsan elbomlanak és oxidálódnak, emiatt ioncsatorna gátló képességüket (azaz biológiai aktivitásukat) elveszítik. Ezen felül a peptidek bőrön keresztül nem szívódnak fel, a gyomor-bél traktusban az emésztőenzimek a peptideket elbontják, csak parenterálisan adhatók be hatásosan a célszervezetbe (kísérleti állatba). A termelt ioncsatorna gátló toxinok környezetre kifejtett hatása elhanyagolható, nem jelent veszélyt.
Rovarsejt Expressziós Rendszerek
A rovarsejt-expressziós rendszerek, jellemzően a Baculovírus expressziós rendszer (BEVS) alapúak, ideálisak nagy, komplex fehérjék, különösen glikozilált vagy diszulfid-kötéses fehérjék előállítására. Ezek a rendszerek képesek a helyes fehérje hajtogatásra és a poszttranszlációs módosításokra, amelyek hasonlóak az emlőssejtekben végbemenőekhez, miközben nagyobb hozamot biztosítanak és könnyebben skálázhatók, mint az emlőssejt-rendszerek. A rovarsejtek alkalmasak komplex fehérjék, például glikozilációt, hajtogatást és komplex módosítást igénylő fehérjék előállítására.
Fúziós Fehérjék és Hasítási Stratégiák
Más hordozófehérjék lehetővé teszik, hogy a kívánt fehérjék meghatározott sejtrészekbe irányuljanak, amelyekből különösen könnyen kinyerhetők, ahol különösen stabilak és/vagy ahol tesztcélokra hozzáférhetőek. Végül a hordozófehérjék speciális tulajdonságokkal is rendelkezhetnek, amelyek például affinitáskromatográfiával történő tisztítást tesznek lehetővé. A legtöbb felhasználási területen a fúziós fehérjéket részesítik előnyben, melyek a hordozófehérjét az aminovégen és a kívánt fehérjét a karboxivégen tartalmazzák. De bizonyos esetekben a fordított változat vagy a kívánt fehérje két fúziós partnerrel történő összekapcsolása is kívánatos lehet. Egy ilyen jellegű fúzióból a kívánt peptid szabad formában történő előállításához a kovalensen kötött fúziós partnerek hasítása szükséges. Ez elvileg kémiai vagy biokémiai (enzimatikus) eljárásokkal érhető el.
Az eddig rendelkezésre álló eljárások limitált specifitása azonban a hasítást többnyire korlátozza, mert fontos a kívánt fehérje elnyerése céljából, hogy egy ilyen jellegű hasítás a fúziós partnerek közötti hasadószekvenciában, tehát az átmeneti régióban, de semmi esetre sem továbbmenően magán a kívánt fehérjén belül következzen be. A fúziós fehérjék szekvenciaspecifikus szétválasztására az eddig alkalmazott kémiai eljárások például egy fehérjén belüli metionin aminosavrész bróm-ciánnal történő hasítása és az Asp-Pro aminosavak között savas közegben, hangyasavval történő hasítás. Ezek az eljárások azonban csak akkor alkalmazhatók, ha a kívánt fehérjében a specifikus hasítóhely, eltekintve a fúziós partnerhez vivő átmeneti régiótól, csak egyszer fordul elő.
A fúziós fehérjék hasítására szolgáló biokémiai eljárások leginkább specifikus proteázok felhasználásán alapulnak. Például a tripszin az arginint vagy a lizint követő peptidkötéseket hasítja a fehérjéken belül. A lizin előzetes kémiai módosításával a specifitás növelhető, és ezáltal az arginin specifikus felismerése csökkenthető (redukálható). Egy további, a biotechnológiában alkalmazott proteáz a Clostripain, ez az enzim a peptidkötéseket az arginin és az ezt követő, tetszőleges aminosav között hasítja. Azon enzimes eljárásokról, amelyeket eddig a fúziós fehérjék hasítására alkalmaztak, egy áttekintés olvasható F. A. O. Marsion összefoglalójában (D. M. Glover, E.: DNA cloning III, IRL PRESS Oxford és Washington DC, 1987).
Ismertek olyan proteázok, amelyek egy bizonyos fehérjét nagyon specifikusan hasítanak. Bár az ilyen szelektív proteázok többsége (amelyek például az emberi komplement- és véralvadásrendszerben előfordulnak) a szubsztrát egy meghatározott helyén hasít, a megfelelő hasító régió egy másik fehérjébe (például fúziós fehérjébe) történő átvitele esetén, az ilyen jellegű proteázok rendszerint még sincsenek abban a helyzetben, hogy hasítsanak. A szekvenciaspecifikus proteázok sora, amelyeket eddig a fúziós fehérjék hasítására feltételesen alkalmaztak, jelenleg az Xa faktorral kezdődik. Ez specifikusan hasítja az Ile-Glu-Gly-Arg-X szekvenciát, ahol X egy tetszőleges aminosav. Azonban bebizonyosodott, hogy általánosan a fúziós fehérjék - amelyek a megfelelő hasítószekvenciát az átmeneti régiójukban hordozzák - hasítására ez a proteáz sem alkalmazható. A fúziós fehérjék hatékony hasításának különös jelentősége van a rekombináns fehérjék prokarióta szervezetekben történő előállításánál. Ennek során ugyanis szükséges egy olyan DNS-szekvencia klónozása, amely a tulajdonképpeni DNS-szekvencia előtt kezdő kodonként egy AUG kodont tartalmaz. Ennek következtében prokariótákban, mint például E. coliban egy olyan rekombináns fehérje fejeződik ki, amely az első aminosav helyén (1-es pozícióban) metioninrészt tartalmaz. Sok esetben azonban az szükséges, hogy az 1-es pozícióban metioninmentes rekombináns fehérjéket állítsanak elő.
Az ilyen fehérjék kinyerése például egy metioninspecifikus peptidáz segítségével történhet, amely az N-terminális metionint hasítja. Ez az eljárás azonban nagyon költséges, mivel a lehasadás (mértéke) csak fehérjeszekvenálás által állapítható meg. A 84/02351 számon nyilvánosságra hozott PCT-beli szabadalmi bejelentésből megismerhető egy N-terminális aminosav lehasítása egy fúziós fehérjéről. Az eljárás során egy exopeptidázzal, előnyösen leucin peptidázzal, lépcsőzetes hasítással távolítják el a fehérje aminosavait az N-terminálistól kezdődően egészen az X-Pro szekvenciáig. Ebből a termékből az X-Pro szekvenciát vagy két lépésben, vagy egy egylépéses reakcióban, postprolin-dipeptidál-amino-peptidáz (EC. 3.4.14) alkalmazásával távolítják el. A 020 290 számú európai szabadalomból egy további eljárás ismert a fúziós fehérjék enzimatikus hasítására. Ezen eljárás során a fehérje fúziós részeinek hasításához a kollagenáz enzimet alkalmazzák egy meghatározott felismerőszekvencia segítségével. Majd a fúziós részek aminosavait egy ezt követő, további enzimatikus kezeléssel távolítják el. Megállapították azonban, hogy a kollagenáz és egyéb endopeptidázok is csak csekély specifitással rendelkeznek [lásd Biochim. Biophys. Acta 271, 133-144 (1972)]. Ismert az is, hogy a fehérjék N-terminális fúziós részeinek hasításához a már említett Xa faktort is felhasználják. A hasítási hatékonyság már említett problémája mellett azonban ez az eljárás is mutat további hátrányokat, éspedig azáltal, hogy esetleg a fehérje belső szekvenciái is felismerhetők és hasadnak.
Az IgA-proteáz Alkalmazása a Fúziós Fehérjék Hasításában
Különböző patogén baktériumfajok (mint például a Neisseria gonorrhoea és Neisseria meningititis a Neisseria nemzetségből, vagy mint a Haemophilus influenzáé és H. aegypticus a Haemophilus nemzetségből), amelyek az emberi nyálkahártyán növekszenek, rokon szekvenciájú proteázokat választanak ki, amelyek a humán IgA1-re specifikusak, és ezért összefoglalóan IgA-proteázoknak vagy „Igase”-oknak is nevezik őket. Az immunglobulin IgA1 igen fontos komponense a kiváltott immunválasznak, amelynek az ilyen patogén organizmusok infekcióival szemben kell védelmet nyújtania. [Áttekintés: Komfeld és Plánt, Rév. Infect. Dis., 3, 521-534 (1981)]. Emellett az IgA-proteáz autoproteolízis révén a saját előfehérjéjét is hasítja. Az IgA-proteáz általi hasítóhelyek például a Pro-Ala-Pro, Pro-Pro, Pro-Arg-Pro-Pro, vagy Ala-Pro-Arg-Pro-Pro szekvenciákban találhatók.

Ezt a feladatot géntechnológiai úton oldották meg oly módon, hogy a fúziós fehérjék átmeneti régiójába bevezették a Pro-X-Pro felismerőhelyet, illetve hasítószekvenciát, és ezen hasítószekvenciát specifikusan hasították egy IgA-proteázzal. A találmány tárgya tehát eljárás fúziós fehérjék enzimatikus hasítására és ezen fúziós fehérjék kívánt részének kinyerésére, amely azzal jellemezhető, hogy (1) egy átmeneti régiót - amely a fúziós fehérje két részét egymással összekapcsolja - géntechnológiai eszközökkel úgy módosítunk, hogy ebben az átmeneti régióban egy Y-Pro-X-Pro szekvenciájú IgA-proteáz felismerőhely, illetve hasítószekvencia található, ahol X egy tetszőleges aminosav és Y egy vagy több tetszőleges aminosav lehet. (2) Az (1) lépésben keletkező fúziós fehérjét a felismerőhelyén hasítjuk egy IgA-proteázzal.
Az „IgA-proteáz” fogalmába a találmány szerint mindazon proteázok beletartoznak, amelyek az IgA-t specifikusan hasítják, amelyek például a Rév. Infekt. Dis., 3, 521-539 (1981) irodalmi helyen vannak leírva. Ugyancsak alkalmasak a rekombináns IgA-proteázok is, amelyeket például a 36 22 221 NSZK-beli közrebocsátási iratban, a Proc. Natl. Acad. Sci. 79, 7881-7885 (1982); Proc. Natl. Acad. Sci. irodalmi helyeken találhatóak. A fúziós fehérjék átmeneti régiójának módosítása a találmány szerinti eljárással előnyösen úgy valósítható meg, hogy a fúziós fehérjék átmeneti régiójába beépítünk egy DNS-szekvenciát, amely az IgA-proteáz-felismerő helyet vagy annak egy részét kódolja, ahol ez a DNS-szekvencia előtt és/vagy után egy vagy több, a fehérjefúzió kívánt részeit kódoló DNS-szakasz van beépítve.
A találmány szerinti eljárásban az IgA-proteáz-felismerő hely az Y-Pro-X-Pro konszenzus aminosavszekvenciával rendelkezik, ahol X egy tetszőleges aminosavat és Y egy vagy több tetszőleges aminosavat jelent. X jelentése előnyösen szerin, treonin vagy alanin. A találmány szerinti eljárás tehát magában foglalja a legalább Pro-X-Pro konszenzus hasítószekvenciájú IgA-proteáz-felismerő hely bevezetését egy tetszőleges fúziós fehérje átmeneti régiójába, például a hordozó fehérje és a kívánt fehérje közé, majd a hasítást és a kívánt fehérje kinyerését egy IgA-proteáz felhasználása által. A Pro-X-Pro hasítószekvenciában X helyén előnyösen Ser, Ala vagy Thr aminosav állhat. Például: a) Pro-Ala-Pro, b) Pro-Pro, c) Pro-Arg-Pro-Pro, d) Pro-Pro, e) Ala-Pro-Arg-Pro-Pro, f) Pro-Ala-Pro-Arg-Pro-Pro.
Amennyiben a találmány szerinti eljárást olyan fúziós fehérjék hasítására alkalmazzuk, amelyekben a kívánt fehérje a hordozó fehérje után következik, az IgA-proteázzal történő hasítás során olyan fehérje keletkezik, amelynek aminoterminálisa az X-Pro szekvenciával jellemezhető. Ez a szekvencia lehet a kívánt fehérje része előtagként, utótagként. Ez a szekvencia általában akkor szerepel előtagként, ha a géntechnológiai úton előállított kívánt fehérje természetes formájában is tartalmazza az aminoterminálisán az X-Pro-nak megfelelő mindkét aminosavat.
A találmány szerinti eljárás minden más, a fúziós fehérjék hasítására szolgáló ismert eljárással szemben azon előnyökkel rendelkezik, hogy meglepő módon általánosan alkalmazható minden olyan fúziós fehérjére, amelyek a megadott hasítószekvenciát az átmeneti régiójukban hordozzák, és hogy az oldhatatlan, oldott, membránasszociált és a sejthez kötött fehérjefúzióknál is alkalmazható. Különös előnye az eljárásnak ezen túlmenően, hogy lehetőséget nyújt olyan fúziós fehérjék, illetve fehérjefúziók hasítására, amelyek a mikroorganizmusokban precipitátumok formájában keletkeznek, és amelyekből azok - mint ilyenek - könnyen dúsíthatók.
Az „Igase” számára szükséges hasítószekvencia beépítése a fehérjefúzió átmeneti régiójába géntechnológiai eszközökkel kivitelezhető. Így például a nukleotidoknak egy sorrendje, illetve egy nukleotidszekvencia, amely a hasítószekvenciát vagy annak egy részét kódolja, kémiai úton szintetizálható, és egy kívánt fehérje DNS-szakaszai közé ismert géntechnológiai eszközök segítségével beépíthető. A nukleotidok egy természetes sorrendje is ennek megfelelően, amely az alkalmas hasítószekvenciát vagy annak egy részét kódolja, beépíthető. A fehérjefúziót kódoló gén előnyösen egy alkalmas (előnyösen indukálható) expressziós szignálszekvencia kontrollja alatt áll, úgy, hogy a követelményeknek megfelelő fúziósfehérje-termék fejeződik ki. A fehérjefúziók termelésére szolgáló gazdasejtekként alkalmas prokarióta és eukarióta (növényi, valamint állati) sejtek felhasználhatók, azonban sejtmentes rendszerek is alkalmazhatók. Az eljárás során alkalmazott hordozófehérjék bármilyen funkciójúak lehetnek, attól függően, milyen tulajdonságokat kell a fehérjefúzióknak kölcsönözniük, mint például meghatározott transzport funkciók, vagy funkciók, amelyek a fehérjefúziók tisztítását vagy azok stabilitását megjavítják, és még sok egyéb. A találmány szerinti eljárás preparatív, valamint analitikai célokra is felhasználható.
A találmány szerinti eljárás különösen előnyösen alkalmazható olyan rekombináns fehérjék, illetve peptidek előállítására, amelyek az N-terminálison metioninmaradékot nem tartalmaznak, olyan fúziós fehérjékből, illetve peptidekből, amelyek a Met-Y-Pro-X-Pro-A aminosavszekvenciával rendelkeznek, ahol X helyén egy tetszés szerinti aminosav, előnyösen Thr, Ala vagy Ser áll, Y pedig egy vagy több tetszőleges aminosavat jelent, amely előnyösen, ha X jelentése Thr vagy Ala, Pro-val végződik, vagy ha X jelentése Ser, akkor Pro-Ala vagy Pro-Pro szekvenciával végződik, és A egy tetszés szerinti aminosavszekvenciát jelent. Az eljárás lépései a következők:1) egy prokarióta sejtnek egy olyan génnel való transzformálását, amely egy Met-Y-Pro-X-Pro-A aminosavszekvenciájú fúziós fehérjét kódol;2) a fúziós fehérje expresszióját a prokarióta sejtben;3) a Met-Y-Pro-X-Pro-A fúziós fehérjének az N-terminális metionintól és/vagy az Y hordozó résztől való elválasztását, IgA-proteáz alkalmazásával.
A fúziós fehérje Y hordozó része egy legalább 1, előnyösen 1-100, különösen 1-50 tagszámú aminosavszekvenciát jelent, amely egy IgA-proteáz által felismerhető hasítószekvenciával végződik. Ha X helyén Ser áll, akkor Y előnyösen a Pro-Ala szekvenciával vagy Pro-val végződik. Egy különösen előnyös kivitelezési mód esetén az Y helyén legalább 5 aminosav áll, amelyek a Pro-Ala-Pro-Arg-Pro szekvenciával végződnek. Az Y hordozórész emellett még további, előnyösen 100, még előnyösebben 50-ig terjedő tetszőleges aminosavat tartalmaz. Erre előnyösen olyan aminosavszekvenciák alkalmasak, amelyek a DNS-szintjén a Met-Y-Pro-X-Pro-A protein expresszióját javítják. A Met-Y-Pro-X-Pro-A protein expressziója DNS-szinten például a béta-galaktozidáz gén fragmenseinek fúziója révén javítható, azaz az Y hordozórész felöleli a béta-galaktozidáz protein egy részét. Más lehetőségek a Met-Y-Pro-X-Pro-A protein expressziójának növelésére a szakember számára ismertek.
A találmány tárgya továbbá egy fúziós fehérje, amely több polipeptidrészt tartalmaz, és amely a különböző polipeptidrészek közötti legalább egy átmeneti régióba beépített egy vagy több Pro-X-Pro szekvenciájú IgA-proteáz-felismerő hellyel rendelkezik, ahol X jelentése egy tetszőleges aminosav, előnyösen azonban Ser, Thr vagy Ala. Példák: a) Pro-Ala-Pro, b) Pro-Pro, c) Pro-Arg-Pro-Pro, d) Pro-Pro, e) Ala-Pro-Arg-Pro-Pro, f) Pro-Ala-Pro-Arg-Pro-Pro, és Thr-Pro.
A jelen találmány magában foglal különösképpen egy fehérjét, illetve peptidet is, amely a Met-Y-Pro-X-Pro-A aminosavszekvenciával jellemezhető, ahol X jelentésére előnyösen Thr, Ala vagy Ser áll, Y egy vagy több tetszőleges aminosavat jelent, és előnyösen, ha X jelentése Thr vagy Ala, Pro-val végződik, ha X jelentése Ser, akkor Pro-Ala szekvenciával vagy Pro-val végződik, és A egy tetszőleges aminosavszekvenciát jelent.
Gyakorlati Alkalmazások és Biztonsági Megfontolások
Az Alpha Lifetech rekombináns fehérje előállítási szolgáltatásokat nyújt, kiváló minőségű rekombináns fehérje előállítási megoldásokat kínálva ügyfeleinek, lefedve a teljes folyamatot a génklónozástól az expressziós rendszer kiválasztásán át a fehérjetisztításig és a minőségellenőrzésig. Elkötelezettek amellett, hogy testreszabott rekombináns fehérje szolgáltatásokat nyújtsanak biogyógyszerek, tudományos kutatóintézetek és gyógyszeripari vállalatok számára, hogy kielégítsék az ügyfelek különböző kutatási szakaszokban lévő igényeit.
A rekombináns fehérjetermelés során felmerülő kihívások közé tartozik az alacsony expresszió, a fehérje nem megfelelő feltekeredése, ami zárványtestek kialakulásához vezet, a nehézségek a fehérjetisztításban, a fehérjelebontás vagy inaktiváció, valamint a fehérje biológiai aktivitásának hiánya.
A Debreceni Egyetem különböző intézeteiben számos kutatási tevékenység zajlik géntechnológiával módosított szervezetekkel és rekombináns fehérjékkel. Az Élettani Intézetben rekombináns fehérjék, illetve azok pontmutánsainak előállítása történik E. coli alapú expressziós rendszerben a fehérje biológiai funkciójának vizsgálatára. A Molekuláris Biológiai Intézetben a sejthalál biokémiai mechanizmusait, a sejthalál és fagocitózis szignálútvonalait, a veleszületett immunitás és a vírusfertőzések kapcsolatait, valamint retrovírusok biokémiáját, transzkripciós folyamatok genetikai és epigenetikai szabályozását, biomarkerek hasznosságát, valamint őssejtek differenciációját kutatják. A Proteomika Szolgáltató Laboratórium célja elsősorban biológiai minták proteomikai és metabolomikai elemzése egyetemek és biotechnológiai cégek számára a proteomika területén.

Az Orvosi Mikrobiológiai Intézetben a GMO-kat érintő tevékenységek magukban foglalják humánpatogén vírusok patogenezisben fontos fehérjéinek, szabályozó régióinak vizsgálatát, a vírus-gazdasejt interakciók tanulmányozását, a különböző virális gének gazdasejtek génexpressziójára gyakorolt hatásának vizsgálatát, potenciális célsejtek keresését; virális antigének előállítását szeroepidemiológiai vizsgálatokhoz; vírusgenomok transzfekciójával 2. kockázati csoportba tartozó, potenciálisan humánpatogén polyomavírusok létrehozását, a replikáció tanulmányozását; a virális nukleinsav kópiaszám meghatározását qPCR-rel lehetővé tevő plazmidok létrehozását.
Az állatkísérletek során génmódosított állatokat, mint például a knock-out (KO) egereket, használnak a specifikus gének célzott tanulmányozására, különösen a betegségek kialakulásában és a terápiás válaszokban. A génmódosítás során transzgenikus technológiákat és homológ rekombinációt alkalmaznak emberi betegségmodellek létrehozására. Az Anatómiai, Szövet- és Fejlődéstani Intézetben a kísérletek célja az agyi fejlődésbeli rendellenességek vizsgálata transzgenikus technológiák kombinatív felhasználásával, valamint a gerincvelői fájdalomfeldolgozó neuronhálózat kialakulásának és működésének vizsgálata ép és kóros körülmények között. Ehhez új génmódosított szervezetek kerülnek létrehozásra in utero elektroporációval vagy tesztikuláris mikroinjekcióval, amelyhez génszerkesztésre alkalmas plazmidokat vagy in vitro transzkripcióval létrehozott RNS szekvenciák, bizonyos esetekben pedig AAV vírusvektorok kerülnek felhasználásra.
Környezetvédelmi és Biztonsági Előírások
A genetikailag módosított mikroorganizmusokkal és állatokkal végzett munka szigorú biztonsági előírásokhoz kötött. A GM egértörzsekben a genetikai módosítások csökkent életképességhez vezetnek, és nem jelentenek környezeti veszélyt. A GM egérvonalak és patkányok környezetbe való kijutásának kockázata az állatházi tartási és az alkalmazott kísérleti körülmények között elhanyagolható. A szállítás lezárt dobozban történik, a dobozok kinyitása csak a laborban zárt ajtók mellett engedélyezett. Amennyiben a GM egerek vagy patkányok az intézkedések ellenére kiszabadulnának, a személyzetnek mindent meg kell tennie a befogás érdekében. A GM állatok nem rendelkeznek emberre, állatra, illetve növényekre veszélyt, s bár szaporodhatnának a vad típusú egerekkel, a genetikai módosítások elterjedése nagyon valószínűtlen. A GM és a GM Hv1 KO egértörzsekben a genetikai módosítások csökkent életképességet eredményeznek, nem laboratóriumi körülmények között fennmaradásuk nem valószínű.
Az expressziós vektorok előállítására használt E. coli DH5alpha baktériumtörzs antibiotikumrezisztenciát tartalmaz. A baktériumok nem megfelelő kezelése az antibiotikumrezisztencia terjedését eredményezheti. A környezeti kockázat szintje alacsony. A laboratóriumban alkalmazott retro- és lentivírus alapú GMV-k kizárólag a transzdukció céljából előkészített eukarióta sejtekbe képesek bejuttatni genetikai anyagot, specifikus sejt-tropizmus mellett (például T-sejtek, tumorsejtek). Az AAV-k képesek emlős sejtekbe jutni, de egyikfajta vektor sem rendelkezik replikációs képességgel, így nem képes új víruspartikulumokat létrehozni, és nem tudja a genetikai anyagot továbbadni más sejteknek vagy szervezeteknek. Ezért a GMV és más szervezet közötti horizontális génátadás csak elméleti szinten lehetséges, gyakorlatilag viszont a használt rendszerek mellett nagyon alacsony valószínűségű esemény, amely megfelelő biztonsági intézkedések mellett elhanyagolható valószínűségű.
A plazmid vektorok amplifikációjára használt kompetens E. coli törzsek a vektor által kódolt antibiotikum-rezisztencia géneket hordoznak. Arra, hogy ezek ne kerülhessenek a környezetbe fokozott figyelmet fordít a Kérelmező. A tevékenység során a laboratóriumban alkalmazott E. coli törzsek molekuláris biológiai munkára lettek optimalizálva, ezáltal a környezetben való túlélésük rendkívül korlátozott. Annak esélye, hogy más, rokon mikroorganizmusokkal képesek kapcsolatot létesíteni, vagy kiszorítani azokat gyakorlatilag elhanyagolható. Semmilyen élőlényre nézve nem patogén szerotípusok, így állatokra, növényekre, gombákra, egyéb élőlényekre való káros hatásuk kifejtése nem várható.
tags: #feherje #expressios #rendszerek #e #coli #eleszto